








 26 / 928
26 / 928

1ГГИТil состояние 2
5.5 -э
5.0 :
to
4.5 -
5^4.0
TVniJIMIMIMM
40
50
50
/о
во
суток
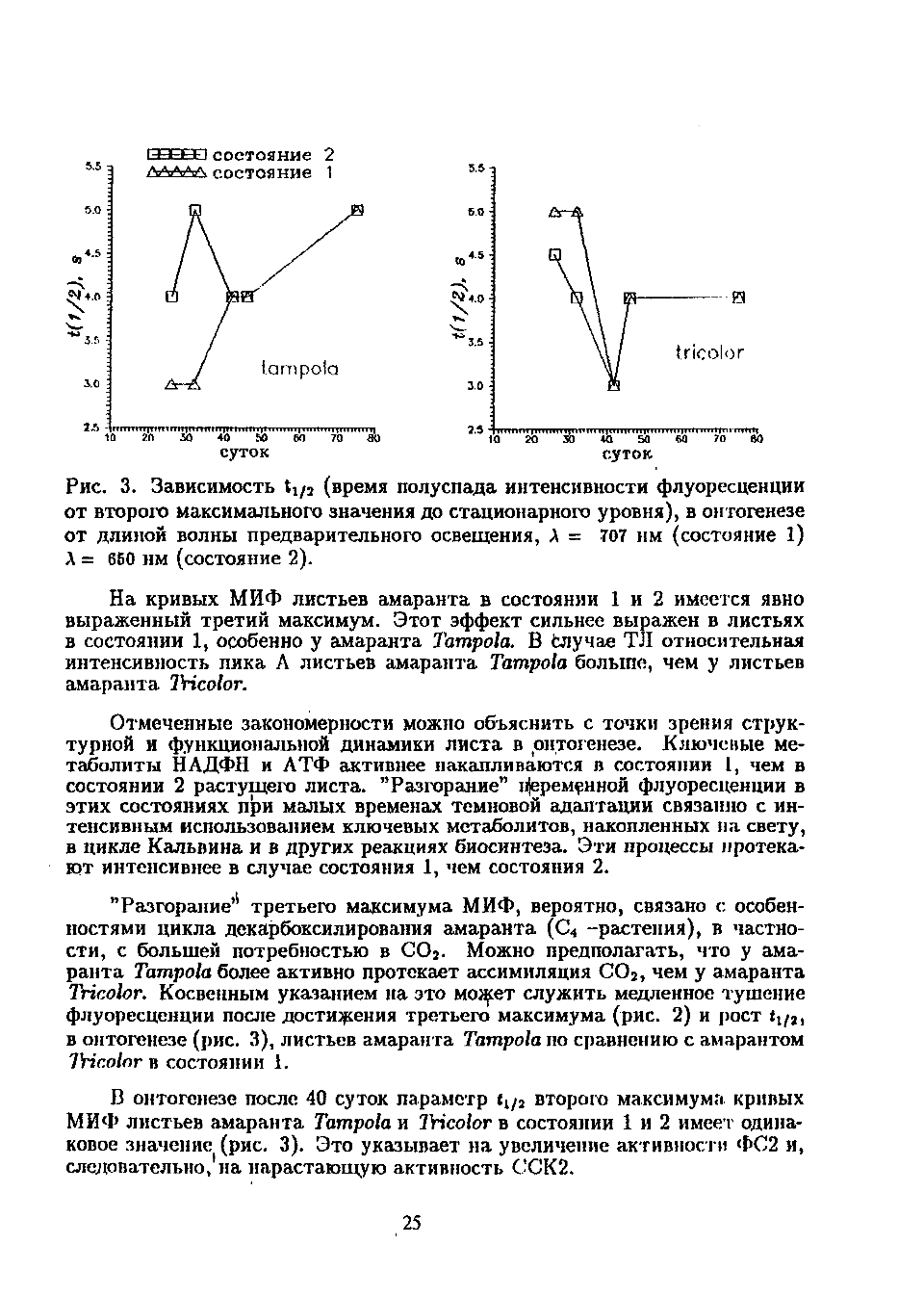
Рис. 3. Зависимость t1/2 (время полуспада интенсивности флуоресценции
от второго максимального значения до стационарного уровня), в онтогенезе
от длиной волны предварительного освещения, λ
=
707
нм (состояние 1)
λ
= 650
нм (состояние 2).
На кривых МИФ листьев амаранта в состоянии 1 и 2 имеется явно
выраженный третий максимум. Этот эффект сильнее выражен в листьях
в состоянии 1, особенно у амаранта
Tampola
. В случае TЛ относительная
интенсивность лика А листьев амаранта
Tampola
больше, чем у листьев
амаранта
Tricolor.
Отмеченные закономерности можно объяснить с точки зрения струк
турной и функциональной динамики листа в онтогенезе. Ключевые ме
таболиты НАДФН и АТФ активнее накапливаются в состоянии 1, чем в
состоянии 2 растущего листа. "Разгорание” переменной флуоресценции в
этих состояниях при малых временах темновой адаптации связанно с ин
тенсивным использованием ключевых метаболитов, накопленных на свету,
в цикле Кальвина и в других реакциях биосинтеза. Эти процессы протека
ют интенсивнее в случае состояния 1, чем состояния 2.
”Разгорание” третьего максимума МИФ, вероятно, связано с особен
ностями цикла декарбоксилирования амаранта (С4 -растения), в частно
сти, с большей потребностью в СО2. Можно предполагать, что у ама
ранта Tampola более активно протекает ассимиляция СО2, чем у амаранта
Tricolor
. Косвенным указанием на это может служить медленное тушение
флуоресценции после достижения третьего максимума (рис. 2) и рост t1/2,
в онтогенезе (рис. 3), листьев амаранта
Tampola
по сравнению с амарантом
Tr
icolor
в состоянии 1.
В онтогенезе после 40 суток параметр t1/2 второго максимума кривых
МИФ листьев амаранта
Tampola
и
Tricolor
в состоянии 1 и 2 имеет одина
ковое значение (рис. 3). Это указывает на увеличение активности ФС2 и,
следовательно, на нарастающую активность ССК2.
25
Научная электронная библиотека ЦНСХБ