






 342 / 540
342 / 540

клеток корней галофита
S. altissima L.
функционирует протонная помпа, Н*-
АТФаза, сходная по своим свойствам с Н*-АТФазой ПМ других растений.
Показано, что внесение Na+в среду вызывает диссипацию сформированного
ДрН на везикулах ПМ. Это было интерпретировано, как NaVH^ обмен,
осуществляемый Na+/H* антипортером [Popova, Bainokin, 1992]. Однако в наших
экспериментах внесение соли Na+ в суспензию везикул ПМ &
altissima
после
генерации ДрН не приводило к разрядке
РГ-градиента, свидетельствуя, что нам не
удалось зафиксировать активность NaVH*
антипортера в полученной мембранной
фракции. Также не обнаружено АТФ-
зависимую генерацию Ду на ПМ, когда
среда была слабо щелочной (pH 8,0).
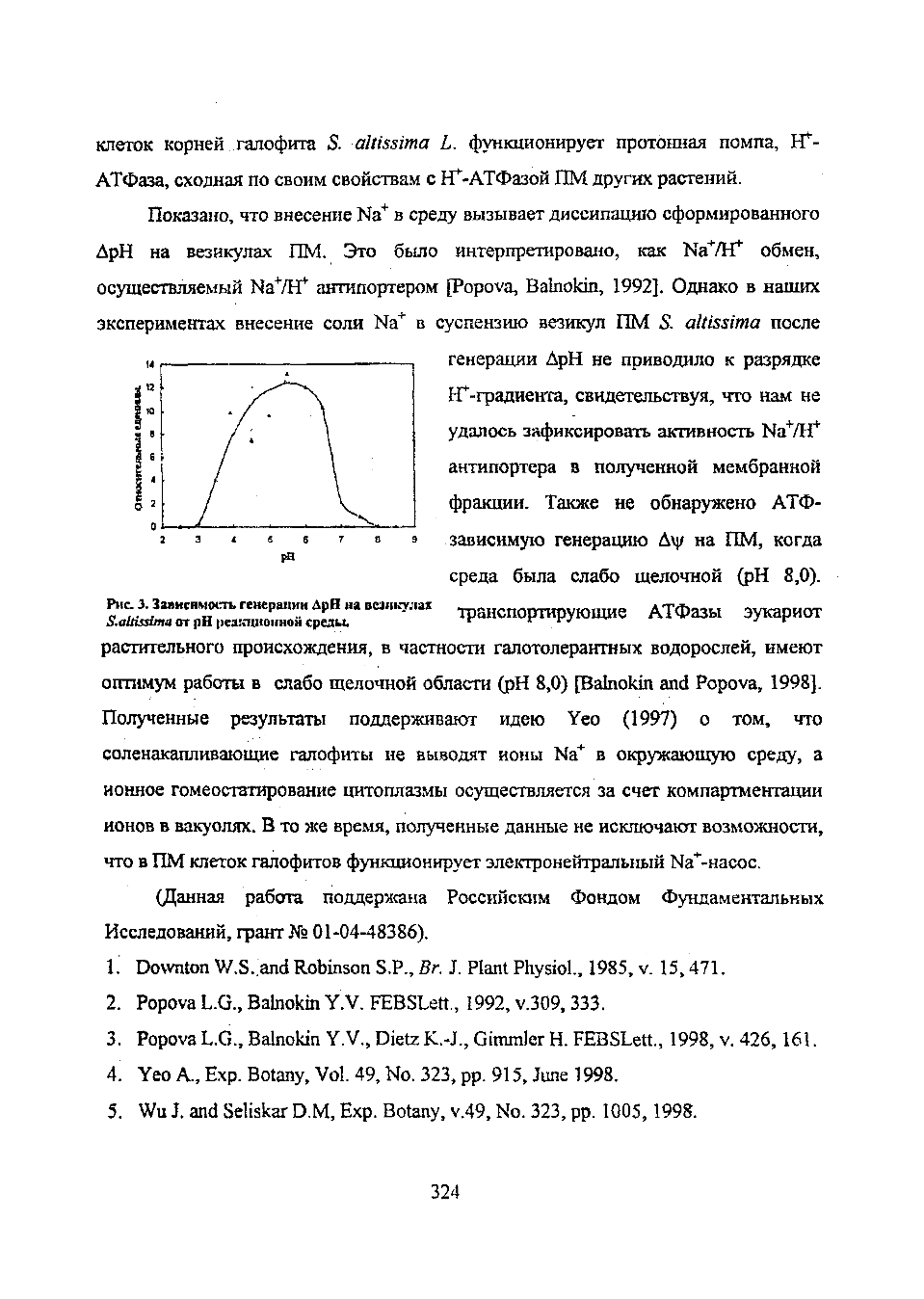
^ Г “ а = Г с р ^ : ИВМНКУЛ“ транспортирующие АТФазы эукариот
растительного происхождения, в частности галотолерантных водорослей, имеют
оптимум работы в слабо щелочной области (pH 8,0) [Bainokin and Popova, 1998].
Полученные результаты поддерживают идею Yeo (1997) о том, что
соленакапливающие галофиты не выводят ионы Na+ в окружающую среду, а
ионное гомеостатирование цитоплазмы осуществляется за счет компартментации
ионов в вакуолях. В то же время, полученные данные не исключают возможности,
что в ПМ клеток галофитов функционирует электронейтральный Na+-Hacoc.
(Данная работа поддержана Российским Фондом Фундаментальных
Исследований, грант № 01-04-48386).
1. Dovvnton W.S. and Robinson S.P.,
Br.
J. Plant Physiol., 1985, v. 15,471.
2. Popova L.G., Bainokin Y.V. FEBSLett., 1992, v.309, 333.
3. Popova L.G., Bainokin Y.V., Dietz K.-JL, Gimmler H. FEBSLett., 1998, v. 426, 161.
4. Yeo
A.,
Exp. Botany, Vol. 49, No. 323, pp. 915, June 1998.
5. Wu J. and Seliskar D.M, Exp. Botany, v.49, No. 323, pp. 1005,1998.
pH
324
Электронная Научная СельскоХ зяйственная Библиотека